Buddleja forrestii and
Buddleja macrostachya
Updated May 2025

This plant of unknown origin was labelled B. macrostachya. On close examination, it turned out to be B. forrestii.
Open full size image in a new window
B. forrestii and B.macrostachya are two closely allied Himalayan species with a significant overlap in range. They both originate from south-western China (notably Yunnan and Sichuan), Bhutan, northern Myanmar, and north-east India, with B. macrostachya ranging further south and east into Bangladesh, Thailand and northern Vietnam. Some examples of these species can be hard to correctly identify, and the species easily confused with one another.
Generally, both grow as large woody shrubs in hillside scrub and in forest clearings, and are usually found at altitudes greater than 1,000 metres. Evolving for a montane habitat makes them more able to cope with adverse weather conditions than one might expect. Different collections vary in their hardiness, but mostly these plants should be able to withstand an average winter in lowland southern England (USDA zone 8), perhaps requiring extra shelter further north.
The taxonomy of these species is not without controversy. The famous plant hunters, such as George Forrest and Francis Kingdon-Ward, sent numerous specimens from the Himalayan region back to UK botanical institutions (such as the Royal Botanic Gardens, Edinburgh) during the early 20th century. This was era of 'splitting' plants into separate species based on very small differences, and as a result a large number of supposedly 'new' species were described. Later examination of both dried herbarium specimens and plants in living collections revealed that many of these specimens could be 'lumped' into just these two species. Therefore, the last systematic review of the old world Buddleja (see Leeuwenberg 1979) sunk a number of these names under B. forrestii and B.macrostachya, reducing many former species to synonyms without any further division into subspecies or varieties. However, it is possible this 'lumping' went too far, with some forms meriting at least varietal status, if not reinstating as a distinct species.

The two species share similar foliage, large leaf blades, which are lanceolate to elliptic in shape with an acuminate (pointed) apex. This is a B. forrestii.
Open full size image in a new window
From my experience of growing several different forms of each species, being able to closely examine the plants, and the results of several crossing experiments, I can see the reasons for reducing many species names to synonyms. Two specimens that may appear rather different can share significantly more similarity on closer examination. However, it is not clear-cut and there might be some justification for reinstating some species.
Identifiction of these species can be confusing: two specimens from different species may be superficially more similar to one another than to members of their own species. The tendency to self-fertility and localised populations of a single type, and the opposite propensity for hybridisation, and hence the possibility of intermediates between the two species, complicate matters still further. However, I hope to show below that this can be untangled and these two species can be reliably distinguished.
Their horticultural merit may be somewhat debatable, as they require space and tend to a rangy, messy habit. However, these species offer the gardener something more exotic than the familiar cultivars, whilst being only a little more challenging to grow. The requirements for both are similar: full sun is best, but because they evolved at altitude they don’t like to be too hot; they need a humus-rich soil which is moisture retentive in summer and well-drained in winter, and they're not fussy about pH. They should be sheltered from the harshest winds and and protected from the hardest frosts. The roots will benefit from a winter mulch which protects the soil from freezing solid. Plants may be killed almost to the ground in really harsh weather, but will regenerate from the hardwood if the root-ball is kept alive.
Buddleja forrestii Diels

The long flower panicle of a B.
forrestii. This plant's native origin is unknown.
Open full size image in a new window
Synonymous species:
Buddleja cooperi W.W.Smith; Buddleja forrestii var.
gracilis Lingelsheim; Buddleja henryi Rehder & E.H.Wilson; Buddleja henryi var. glabrescens C.Marquand;
Buddleja latiflora S.Y.Pao; Buddleja limitanea W.W.Smith; Buddleja longifolia Gagnepain; Buddleja pterocaulis
A.B.Jackson; Buddleja subherbacea Keenan; Buddleja
taliensis W.W.Smith.
B. forrestii is a highly variable Himalayan species and is closely allied with B. macrostachya. There are quite a few individual cultivars (or collections) around the UK in various botanical gardens and plant collections. Sometimes their origins have been lost or forgotten, and in some instances even labelled as an entirely different species.
This plant is from Roseland House Garden and Nursery in Cornwall, and I’ve tracked it back to Forde Abbey in Dorset. Beyond that, the trail goes cold. It has many typical features of the species, such as large robust leaves and four-angled winged branchlets. The inflorescences, which appear in July and August, are long and pendulous. The flowers are just off-white, sometimes with a pinkish cast, emerging from mauve-flushed buds.
I’ve found this example of the species to be reasonably hardy, down to at least -10ºC, and these large species Buddleja do become hardier with size and age. It’s proved semi-evergreen, although the foliage suffers in the cold, and it may even be killed back to the old wood in very harsh winters. The roots should survive, producing strong new growth as the spring warms up. Already about 3 metres in height and spread after just a few years, I expect the plant to get bigger, maybe even reaching the proportions of a small tree.
The white-flowered plant shown at the top of the page, which came to me labelled up as B. macrostachya, is quite similar, except overall it's much less vigorous and fully deciduous most winters. Its origins are lost and set to remain a mystery.

W/O 7061 from the south of Sichuan.
Open full size image in a new window
W/O 7061 comes from seed collected in south Sichuan (Liangshan at 2775 metres) by Chinese Alpines, and I have several siblings. At first, they were pretty much alike, but have later developed a number of variations. The one shown has a strong mauve flush to the buds and the flowers become slightly yellow as they mature.
Some of the siblings have pure white flowers, such as the one which is shown on the Supplementary Gallery page.
A couple of plants of the collection W/O 7061 I've raised show features associated with B. macrostachya (tomentose exterior to the corolla, corolla tube longer than 6mm) and the identification as B. forrestii is possibly ambiguous. However, taking together all the features they are within the natural variation of the species, and I do not believe they are hybrids or intermediate forms between the two species. Certainly, I’ve found the two species relatively reluctant to cross in my own garden, although several of the various B. forrestii types have been successfully pollinated by B.davidii and B. nivea.
It may be coincidental that all the examples of B. forrestii (excluding the limitanea type, see below) I’m growing have predominantly white flowers, as the literature suggests the species may have violet, mauve, purple or even wine-red flowers. However, I've yet to find a strongly coloured example of this type or even seen a photograph of a such a plant. Herbarium specimens of the synonymous B. taliensis describe the flowers as 'maroon-crimson', but no living specimen of this type is known to be in cultivation.

B. pterocaulis should be a form of B. forrestii.
Open full size image in a new window
Buddleja forrestii
type pterocaulis
Several species were sunk under B. forrestii by Leeuwenberg (1979) including B. limitanea W.W.Smith (see below) and B. pterocaulis A.B.Jackson (originally called B. longifolia Gagnep., which has caused some confusion). Plants labelled 'B. pterocaulis longifolia', which is not a legitimate name, are supposedly a form of B. forrestii. It would seem extant ex situ examples are derived from a specimen once grown at Borde Hill, West Sussex (see Marquand 1930 and Jackson 1935).
The name pterocaulis references the winged stems, although the wings of this form are no more conspicuous than any other form. No one I’ve contacted seems to remember seeing the flowers, reportedly a purple- or violet-colour. I have managed to persuade my pot-grown specimen to flower. Although the leaves and branchlets are somewhat more pubescent than average for the species, the flowers are absolutely typical - mostly glabrous ovary and exterior to corolla, stigma level with the stamens and funnel-shaped - and they have proved to be white rather than violet.
I've found another source for this type, and I'm hoping this second 'pterocaulis' will have the reported violet-hued flowers.

ACE 2522 is a Buddleja forrestii of the type previously described as B. limitanea.
Open full size image in a new window
Buddleja forrestii
type limitanea
A Chinese Buddleja which was once treated as a species in its own right, but is now considered a smaller type of B. forrestii. However, it is still frequently labelled as B. limitanea in horticulture, so here it is described as a type, rather than a subspecies or variety. Typically, it has pendulous panicles of mauve-purple to white flowers that appear in mid- to late-summer and may continue to flower until well into the autumn. The individual flowers are larger than B. davidii and have the typical short, funnel-shape corolla of B. forrestii; they can be purple in bud, but often fade after they open. The leaves are usually smaller than the larger forms of B. forrestii, and sometimes slightly grey-felted. Less vigorous than other forms of B. forrestii, it grows to about three metres height and spread. It’s quite hardy, but can be cut back hard in cold winters. If pruning is required, and it can become quite straggly, it should be done in early spring like B. davidii.
There are several different cultivars (or 'collections') around the UK. Buddleja sp. ACE 2522, collected by the Alpine Garden Society, KR 2737, collected by Keith Rushworth, and BO 14-138, collected by Bjørnar Olsen are all from the Cangshan in Yunnan, China. All these are quite similar, although the flower colour varies from whitish flushed with mauve to pale pink to a deeper mauve.
There are also cultivars of unknown origin. One, which has lost its history, has been erroneously labelled as B. delavayi, a species totally unlike any B. forrestii. This plant is a very attractive form, with large individual flowers of a soft pink-lilac, and paler green foliage. Reputedly less hardy than others of the type, I’ve found it quite forgiving. See the Supplementary Gallery page.
There is also a plant labelled as B. limitanea, which is somewhat more ambiguous and whose origins are unknown. In most details, it looks much the same, except for the flowers: these are mauve-purple and have a long corolla tube; the flower parts also have a pubescence not common in forms of B. forrestii. I suspect this plant may be hybrid in origin, either natural or more likely raised ex situ. Just possibly, it is Buddleja X ‘Hotblackiana’, a hybrid of B. forrestii and B. davdii once listed by the RHS and now thought lost. The flower morphology is similar to a B. forrestii X B. davidii hybrid I raised myself. See the Supplementary Gallery page for more details.

The limitanea type does not seem to readily cross with other forms of B. forrestii, although it will hybridise with other relatively unrelated species in my garden, notably with B. nivea. The individual flowers might be morphologically very similar to other forms of B. forrestii, but otherwise the type is somewhat dissimilar. Because the limitanea type is recognisable and distinct, I would suggest it as least afforded varietal status as Buddleja forrestii var. limitanea and maybe even returned to the rank of species as Buddleja limitanea W.W.Smith.
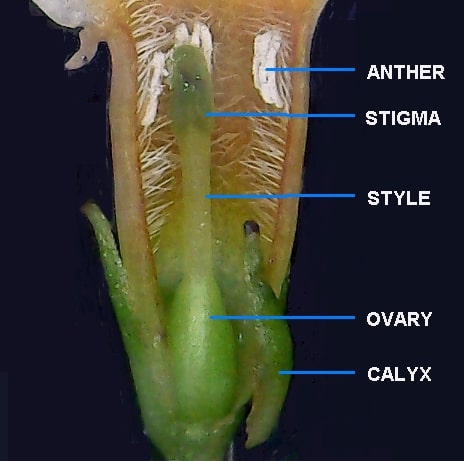
The flower of B. forrestii flares towards the mouth, has anthers level with stigma and the ovary is hairless.
Flower Morphology
Sometimes B. forrestii and B. macrostachya are difficult to tell apart, as the two species can be so similar. Gross features such as panicle form, foliage, stem shape and growth habit cannot be relied upon to reliably identify the species. Only close examination of the flowers can effectively distinguish them. As a general rule, the ovary, style and exterior of the corolla are glabrous (virtually hairless) in B. forrestii, whereas these parts are stellate-tomentose (with a covering of tangled hairs) in B. macrostachya. The corolla tube is short (6-9 mm), wider and flares towards the mouth; in B. macrostachya the tube is 9-14mm long and cylindrical. In both species, the almost sessile anthers are placed close to the mouth of the flower: level with the stigma in B. forrestii and somewhat above in B. macrostachya. The interior of the tube may be pilose (with longer, straight hairs).
Buddleja macrostachya Benth.
Synonymous species:
Buddleja cylindrostachya Kränzlin;
Buddleja hancockii Kränzlin;
Buddleja henryi (Rehder & E.H.Wilson) var. hancockii (Kränzlin) C.Marquand;
Buddleja hookeri C.Marquand;
Buddleja hosseusiana Kränzlin;
Buddleja martii Schmidt.

Buddleja macrostachya RF 040: this particular example shows how closely this species may resemble B. forrestii. Only close examination reveals its correct identifiction.
Open full size image in a new window
B. macrostachya is also a very variable species with a wide range across the Himalayan region and south-east Asia, and frequently co-existing with B. forrestii. Although sympatric, the two species do not appear to frequently hybridise. The flowers of B. macrostachya can be white through to wine-red, the panicles upright or pendulous, and the flowering time can vary from summer to mid-winter. This variation in form led the plant hunters of the late 19th and early 20th centuries to supposedly 'discover' a number of discrete species, although now it is generally accepted that many of these can be reduced to synonyms. There is data to suggest B. macrostachya is a young species and may still be undergoing a rapid evolution and divergence, although this does not suggest the various forms have already divided into truly separate species (Chen at al 2007). This taxon is therefore probably best described as a species complex.
Whilst some types of B. macrostachya are easy identified, others can be virtually indistinguishable from B. forrestii without close examination of the flowers. Some forms may be distinguishable by their felted or glaucous leaves, although this is a far from reliable indicator. There are quite a few different specimens around in the UK, both in the horticultural trade and in botanical gardens.
One plant I’ve grown is, I believe, RF 040 from Arunachal Pradesh, north-east India, and is currently being mislabelled as 'B. forrestii'. It demonstrates how B. macrostachya can so closely resemble B. forrestii in all but the smallest details. This collection forms a large semi-evergreen shrub, with large leaves and rounded branchlets, both covered in an unusual coppery-brown tomentum. RF040 flowers in summer (around early August); the panicles are long, pendulous and many flowered, and loved by the bees. The colour is cream, but can appear slightly pale-pink in some lights. The most obvious features in determining this plant as B. macrostachya are the stellate-tomentose (with tangled hairs) exterior of the long, cylindrical corolla tube and the calyx. This has proved quite hardy (down to at least -5ºC) once established, although much lower than this and it will shed all its leaves and suffer some die-back. After being cut back by frosts, it can be releuctant to flower.

{kind=link}
{kind=link}
SBEC 360 was collected by Roy Lancaster in 1981 on the Sino British Expedition to Cangshan (SBEC), Yunnan, China. Allegedly, his personal specimen was so vigorous it had to be removed before it damaged his house. It grows into a very large shrub that flowers in the summer months, and is likely self-fertile. Identification as B. macrostachya is less certain than the others mentioned, as the ovary is almost glabrous, but the corolla tube is cylindrical and somewhat tomentose, so I think the designation is probably correct. SBEC 360 has been mistakenly labelled as B. x alata in some gardens.

The flowers of SBEC 360 have shorter corolla tubes with sparser tomentum than average for the species, so the identification as B. macrostachya is less certain.

Buddleja sp. GWJ9286, recently given the name 'Sikkim Snow', ia actually a small form of Buddleja macrostachya
Open full size image in a new window
GWJ 9286 was collected in the Lachen Valley, Sikkim (northern India) by Crûg Farm Plants, North Wales. It's been labelled variously as B. paniculata, species nova and B. myriantha v. alba 'Sikkim Snow'.
However, I have positively identified the collection as a form of B. macrostachya, although GWJ 9286 is atypical for the species, being smaller overall: about 2 metres height and spread and with more delicate silvery foliage. The flowers themselves conform precisely to the botanical descriptions of B. macrostachya, although the flower-panicle is more open (interrupted) than others of this species (see Leeuwenberg 1979)
Going back to C.V.B. Marquand's 1930 Revision of the Old World Species of Buddleja, one previously recognised species was Buddleja hookeri, now synonymous with B. macrostachya. Marquand's description of B. hookeri is remarkably similar to GWJ 9286 in many respects - small calyx, tomentose ovary and corolla tube, lax flower panicle and subterete (rounded) stems. Moreover, the original report cites the Lachen Valley, Sikkim as the location of the B. hookeri holotype, the same as GWJ 9286. Leeuwenberg also notes the B. hookeri type as having interrupted panicles. The literature provides strong evidence for GWJ 9286 being this form of B. macrostachya, and, at my suggestion, the plant is currently labelled B. hookeri 'Sikkim Snow’ by Crûg Farm.
Having grown the plant for a few years, I would recommend it as by far the best form of B. macrostachya for the average garden with its fine foliage, restrained habit, and attractive blooms, which are strongly scented as an added bonus. As with RF 040, it can suffer die-back in hard winters.
I’ve also had the opportunity to attempt different pollinations and to grow its progeny. Unlike many of the B. forrestii plants, I’ve found B. macrostachya plants reluctant to hybridise. GWJ 9286 has crossed quite readily only with RF 040; the resulting reciprocal seedlings are intermediate between the types and immediately recognisable as B. macrostachya, leaving me in no doubt as to my identification. See the Supplementary Gallery to view the seedlings.

Buddleja macrostachya PAB 4198, a late flowering collection from India.
Open full size image in a new window
The species is variable with regard to flowering time, and some collections flower much later in the year – in the autumn or even after mid-winter. As far as I can ascertain, this appears to be more common in southern populations; possibly it’s a strategy to avoid coinciding flowering with the monsoons and taking advantage of a longer growing season.
PAB 4198 is a collection by Paul Barney of Edulis Plants, originating from Shillong, (Meghalaya, India) and has fleshy pink flowers in upright panicles. It flowers very late in the season, about November. It needs both a good summer and a mild autumn for it to bloom well in the UK. In my garden, it's formed a small tree and was over four metres in height. Unfortunately, this type of B. macrostachya is not completely hardy and a hard winter killed it back to the ground. The regrowth has never been sufficient for it to flower again.
On casual examination, this example appears very different to RF 040. However, the individual flower morphology is exactly the same. The exterior of the corolla tube and the calyx has the same tomentose appearance, and the ovary and style (pistil) are practically identical. Based on this close similarity, it can be concluded PAB 4198 is the same species as RF 040, and the change in phenology a geographical adaptation. Alternatively, the differences may be an indication of nascent speciation (see below).
{kind=link}
Other interesting winter flowering forms are HWJ 602 and WWJ 12016, collected by Crûg Farm Plants in north Vietnam at the eastern extreme of the species' range. A plant called 'Fansipan', also collected from northern Vietnam, is available commercially in Australia. This has upright panicles with pink-red flowers and closely resembles WWJ 12016.

An example of the species growing in situ at 2000m, Lenteng, Mizoram, India. The late-flowering types have these very
upright and dense panicles. (Photo by Paul Barney, Edulis Plants.)
Open full size image in a new window
A note on Taxonomy
Recently, Chen et al (2021) divided B. macrostachya specimens from Yunnan into two categories: Type 1) those flowering during the warm-wet season (July-September) and with pendulous ('nodding') panicles; and Type 2) flowering during the winter dry season (November-March) and with upright ('erect') inflorescences. There is an evolutionary logic, as downward facing flowers would drain the water away; those flowering in the dry season would need no such adaptation.
The authors also observed that Type 1 has double the chromosome number of Type 2 - in other words, as well as being separated by phenology and morphology, they have diverged with regard to their ploidy too. As an evolving species, it is likely there are intermediate or alternative forms which do not precisely fit the hypothesis.
B. macrostachya is best described as a species complex, but there is an argument that the two types of B. macrostachya are sufficiently divergent to be afforded at least varietal status. As far as I know, the upright Type 2 was the type originally described as B. macrostachya (Bentham 1835). Type 2 has also been reported in the literature as Buddleja cylindrostachya (Kränzlin 1913) and Buddleja hosseusiana (Kränzlin 1923), so these can be reduced to synonyms.
Which leaves the issue of what Type 1 might be renamed if a separate species were reinstated or a varietal name (=subspecies) created. Reference to Marquand (1930) and Leeuwenberg (1979) would indicate Type 1 has been published as B. hancockii (Kränzlin 1913), B. hookeri (Marquand 1930), and Buddleja henryi var. hancockii (Marquand 1930). It should be noted B. hookeri is almost certainly the same species as B. hancockii, differing only in having a more interrupted panicle.
The rules of nomenclature usually give precedence to the first published name. If accepted as two separate species, Type 1 would become B. hancockii, Type 2 staying as B. macrostachya. An alternative would be to make a varietal addition for Type 1: B. macrostachya var. hancockii.
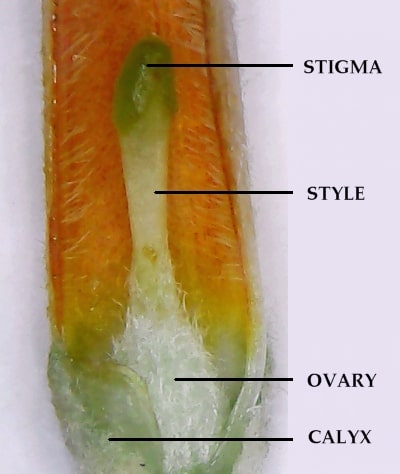
The corolla tube of B. macrostachya is cylindrical, and the ovary and calyx are covered in tangled hairs.
Flower Morphology
Buddleja macrostachya is most easily distinguished from the similar B. forrestii by dissecting the flowers.A B. macrostachya RF 040 flower has been cut open to reveal the pistil and ovary. Note that the ovary (and later the seed capsule) is covered in tangled hairs (stellate-tomentose) and how the base of the style is also hairy. The interior of the corolla tube has numerous hairs (villose) down to level with the ovary and the calyx is tomentose (covered in hairs). In comparison, the flower of B. forrestii lacks this tomentum: the ovary, calyx and exterior of the corolla tube is glabrous or with very few hairs; additionally, the interior of the corolla tube usually has hairs increasing towards the throat. In B. macrostachya, the stamens are in the throat of the flower and held above the stigma, whereas in B. forrestii the stamens are typically level or almost level with the stigma.
Hybrids

This B. forrestii x B. macrostachya hybrid could be either species from a distance.
Open full size image in a new window
A B. forrestii x B. macrostachya hybrid was reported by Leeuwenberg (1979). Possibly, because the two species can be easily confused, this was one or other of the species rather than an actual hybrid. B. x griffithi Marquand has been both reported as a variety of B. macrostachya and as a hybrid of B. macrostachya and B. candida. More details here.
In my garden, I have found B. macrostachya very reluctant to cross with other species, whereas B. forrestii will hybridise with several other species. I have successfully produced only one B. forrestii x B. macrostachya hybrid, using the white flowered B. forrestii shown at the top of the page as the seed parent, and using B. macrostachya RF 040 as the pollen donor. The plant is a perfect intermediate. Dissection of a flower shows the stigma is only slightly below the level of the stamens, whereas in B. macrostachya the stigma is typically much lower due to the long corolla tube, and in B. forrestii the stigma is practically level with the stamens because the tube is relatively short. The tube length itself is intermediate between the parent species.
{kind=link}
It is interesting to speculate whether the large number B. forrestii and B. macrostachya synonyms may be due in part to the collecting and describing of naturally occurring hybrids and intermediates by the plant-hunters, with any taxonomically ambiguous plant receiving a new epithet. It also can't be discounted that hybridisation plays a significant part in the infraspecific diversity of these and other Buddleja species.
Key for distinguishing the species
Both species are highly variable, and features will vary across each species. The feature most useful for a correct identification is the individual flower, with the shape and indumentum of the corolla tube being the most consistent and reliable difference between the species; when considering to which species a specimen should be assigned, this feature should be the most heavily weighted.
The seeds capsules of B. macrostachya remain
covered in hairs and the dried pistil can be persistent.
Open full size image in a new window

| B. forrestii | B. macrostachya | |
| Stems | Branchlets four-angled, winged; pubescent at first, later becoming glabrescent. | Branchlets terete to four-angled, sometimes narrowly winged, white or tan indumentum. |
| Leaves | Leaf underside only thinly tomentose. | Leaf underside densely stellate-tomentose. |
| Corolla tube. | 6-9mm long, funnel-shaped, exterior glabrous or almost glabrous. | 9-14mm long, cylindrical, exterior tomentulose to densely stellate-tomentose. |
| Stamens | Inserted at the mouth, level or almost level with the stigma | Inserted at or near the mouth, somewhat above the stigma. |
| Ovary and base of style | Glabrous to almost glabrous with few hairs, or occasionally loosely tomentose on upper half only. | Lightly pubescent to densely stellate-tomentose. |
| Seed capsule | Glabrous or glabrescent, end rounded without persistent pistil. | Retaining tomentum, end pointed with persistent dried pistil. |
Acknowledgments
For plants, photos and conversations, my thanks to:
Paul Barney - Edulis Plants
Peter Brownless - Royal Botanic Gardens, Edinburgh
Andrew Bullock - The Lavender Garden (now closed)
Nick Lock- Nick Lock Plants
Peter Moore -
Longstock Park
Nursery
Bjørnar Olsen - Chinese Alpines (now closed)
Bleddyn Wynn-Jones - Crûg Farm Plants
References
Chen G., Sun W-B., Sun H. (2007) Ploidy variation in Buddleja L. (Buddlejaceae) in the Sino-Himalayan region and its biogeographical implications. Botanical Journal of the Linnean Society 154:305-312.
Chen, Y., Xu, S., Meng, L., Wang, S, & Chen, Y., Gong W. (2021) Ploidy differentiation and floral scent divergence in Buddleja macrostachya (Scrophulariaceae) complex. Biochemical Systematics and Ecology. 96. 104271.
A.B.Jackson (1935) Buddleja pterocaulis in Journ. Bot. 73:81.
Selected pages from A.J.M. Leeuwenberg (1979) The Loganiaceae of Africa XVIII: Buddleja - Revision of the African and Asian Species: B. forrestii pp124-127; B. macrostachya pp133-136.
Selected entries from Flora of China (1996). 7. BUDDLEJA Linnaeus. 15: 329–337.
C.V.B. Marquand (1930) Revision of the Old World Species of Buddleja. Bulletin of Miscellaneous Information No.5 pp177-208. Royal Botanic Gardens, Kew.
Email:
buddlejagarden@gmail.com
© The Buddleja Garden 2011-2025.